 تويت
تويت
فشل نظرية النشوء التعايشى Endosymbiosis
“simple cells just don’t have the right cellular architecture to evolve into more complex forms”..."Never-ending natural selection, operating on infinite populations of bacteria over millions of years, may never give rise to complexity. Bacteria simply do not have the right architecture."
Chance: The Science and Secrets of Luck, Randomness, and Probability (New Scientist, 2015) p.28-29, 32
يخبرنا بعض العلماء فى كتاب أشرفت على اصداره مؤسسة New Scientist المشهورة بمجلتها العلمية أن أن الخلايا بدائية النوى لا تملك الهيكل المطلوب لتتحول الى حقيقية نوى و أن الانتخاب الطبيعى لو استمر فى العمل على أعداد لانهائية من البكتيريا لملايين السنين فلن يحولها لخلايا أعقد لأن هيكلها لا يسمح بذلك...حسنا هل نعتبر هذا اعترافا بعجز نظرية التطور فى المرحلة الأولى - مجرد خلية تتحول الى خلية أعقد و ليس حتى كائن متعدد الخلايا؟ لا طبعا فسيبدأ الخيال التطورى فى العمل لا يخترع سيناريوهات غير واقعية ليزعم لاحقا أنها حقائق علمية تطورية
يمكن تقسيم جميع الخلايا الى نوعين رئيسيين بدائيات النواة prokaryotes (خلايا بدو عضيات) و حقيقيات النواة eukaryotes (خلايا بها عضيات) و العضيات هى هياكل و أجسام وظيفية موجودة فى قلب الخلية. بين النوعين فجوة تطورية ضخمة خالية من الأشكال الانتقالية و يحاول التطوريون سد هذه الفجوة بوضع فرضياتمنطقية لا بالاعتماد على الأدلة التجريبية
توجد نظريتان طبيعيتان بشأن تكون العضيات هما:
1-النشوء الذاتى autogenous التى تقول بتطور العضيات عن طريق طفرات و انتخاب طبيعى جيل بعد جيل [كالعادة] و أن النموالداخلى للأغشية داخل الخلية ingrowing membranes شكل العضيات
2-فرضية النشوء التعايشى Endosymbiosis و تفترض أن العضيات (مثل الميتوكوندريا و البلاستيدات الخضراء chloroplasts والأسواط flagella ) كانت يوما ما بكتيريا مستقلة تعيش بذاتها ثم ابتلعتها بكتيريا أخرى فتطورت بداخلها و تولت القيام بوظائف متخصصة فى شكل من أشكال التكافل.
ان أول و أكبر دليل على هذه الفرضية العجيبة هو التنادد و هو الدليل التطورى الأثير الذى أكل عليها الدهر و شارب بل و قضى حاجته عليه أيضا. يصر التطورى أنه حال وجود تشابه بين أ و ب فان هذا دليل على انحدارهما من سلف مشترك بينما هذا ببساطة سبببه قيامهما بنفس الوظيفة. مثال بسيط يؤكد لنا التطورى ان وجود دهون Cardiolipin دليل دامغ على انحدارها من البكتيريا التى تملك هذه الدهون لأن حقيقيات النوى لا تملك هذه الدهون فى الغشاء. كالعادة التطورى يتجاهل أن هذه الدهون تستخدم فى تثبيت و دعم و تسهيل عمل lubricating بروتينات توليد الطاقة فى الغشاء لذا تملكها البكتيريا و الميتوكوندريا لأن كلاهما يولدان طاقة من خالال بروتينات مغروسة فى الغشاء بينما لا تملكها حقيقيات النوى لأنها لا تولد طاقة من الغشاء.
Anna Duncan, Alan Robinson, and John Walker, “Cardiolipin Binds Selectively but Transiently to Conserved Lysine Residues in the Rotor of Metazoan ATP Synthases,” Proceedings of the National Academy of Sciences USA 113 (August 2016): 8687–92
Giuseppe Paradies et al., “Functional Role of Cardiolipin in Mitochondrial Bioenergetics,” Biochimica et Biophysica Acta—Bioenergetics 1837
سبب التشابه هو وجود نفس الوظيفة فهو كمن يقول لك أن الهاتف المحمول و الحاسب الالى المحمول يحتويان على الليثيوم بسبب انحدارهما من سلف مشترك بينما هما يحتويان على الليثيوم لأنه مكون فى البطاريات التى بدورها مشتركة ليس بسبب السلف المشترك بل بسبب الوظيفة و قس على هذه الكثير من التشابهات المزعومة التى يستدل بها على أن الميتوكوندريا كانت بكتيريا
يقول السيناريو الشائع أن كيانات كانت يوما مستقلة تضافرت فى مجتمعات تعاونية تكافلية نسميها الخلايا ثم تطورت هذه المجتمعات المتكافلة symbionts الى أكثر من 250 نوع من الخلايا المتخصصة التى تشكل الكائنات متعددة الخلايا فأصبح بعضها عصلات و أخرى عظام أو جلد أو مخ [فى صورة باهرة لأهمية التكافل الاجتماعى] و لكن هذا السيناريو الافتراضى يتطلب مليارات الأشكال الانتقالية غير الموجودة بين أولى بدائيات النوى و المئات من أنواع خلايا الأنسجة المتخصصة كالأعصاب و العضلات و العصى و المخاريطفى الشبكية و غيرها
تفترض النظرية أن الكائن المتكافل كالميتوكوندريا خسر الكثير من جيناته الأصلية أثناء التكيف و ان كان هذا صحيحا فمن المفترض أن الكائنات المختلفة خسرت جينات مختلفة و مع ذلك تتماثل شرائط الميتكوندريا لكل كائن معين هذا الى جانب أن حمض الميتوكوندريا النووى و الرايبوسوم [الة قراءة و ترجمة الحمض النووى] أصغر بكثير من مثيلاتها فى البكتيريا [التى يفترض أنها أصل الكائن المتكافل]
تعتبر هذه الفرضية الأكثر قبولا لأنها الأكثر منطقية [من وجهة نظر التفسير المادى الرافض للتصميم] و ليس لوجود أدلة تجريبية عليها بل انها غير قابلة للاختبار فهى تعتمد فقط على التشابه [حجة التشابه و القوالب النموذجية ideal archetypes تمت مناقشتها فى فصل سابق]
مشاكل الفرضية:
الى جانب كونها مجرد نظرية غير قابلة للاختبار و بلا دليل تجريبى فحتى جانبها النظرى له ما يعارضه فى العضيات الموجودة فعلا مثل:
1-بروتين Tubulin المكون للأنابيب المجهرية لا يوجد فى أى من بدائيات النوى المفترض اندماجها مع خلايا أخرى لتكوين هذه الهياكل
2-السوط flagellum لا يوجد فيه حمض نووى يدعم فرضية أنه كان كائن مستقل يوما ما
3-يستدل الدراونة بتشابه الحمض النووى لبعض العضيات مع بعض بدائيات النوى و لكن من ناحية أخرى هناك تشابهات مع الحمض النووى لحقيقيات النوى تنفى كونه حمض بدائيات نوى فى الأصل
4-لا يوجد سبب يمنع الخلية المضيفة من هضم الكائن المغير عوضا عن استقباله و دمجه فيها
5-تحتاج حقيقيات النوى الى هياكل كثيرة أخرى وظيفية معقدة كثيرة كالأنابيب المجهرية microtubules المهمة لانقسام الخلية و الحركة داخلها و غيرها و التى يجب أن توجد بشكل متزامن لتتفاعل مع بعضها و أقصى ما تفعله النظرية هو محاولة تفسير اثنين من العضيات
الميتوكوندريا:
الحجة الأكثر شيوعا هى اختلاف ال RNA و الرايبوسوم ribosome الخاصة بالمايتكوندريا [وهما من اليات قراءة الحمض النووى و تحويله الى بروتين] عن ما تصنعه النواة لكن هذا ببسساطة يمكن تفسيره بالدور المختلف للمايتوكوندريا...لكن مهلا ما هى الميتوكوندريا أصلا؟
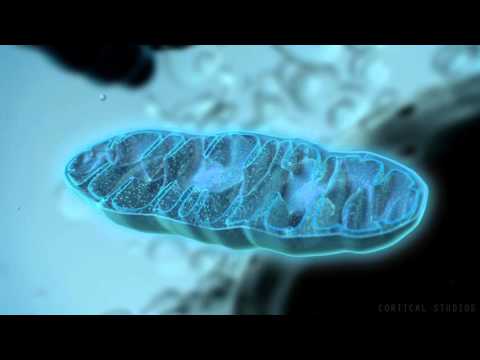
هى هياكل خلوية معقدة هامة تحتوى على اليات و انزيمات تحويل الغذاء الى جزيئات عالية الطاقة ATP وهى مفاعل نوليد الطاقة فى الخلية عن طريق سلسلة من التفاعلات و تحتوى على هياكل تصنيع البروتين كالحمض النووى و الرايبوسوم و لكنها مصغرة و لكن غالبية الجينات المتحكمة فيها فى النواة المركزية للخلية و ليست فى العضيات نفسها مما ينفى كونها كائن مستقل حيث يشفر الحمض النووى للميتوكوندريا نسبة ضئيلة من احتياجاتها بينما تأتى معظم هذه الاحتياجات من تشفير الحمض النووى للخلية المضيفة
Schatz, Gottfried. 1997. "Just Follow the Acid Chain" Nature. 338: 121

لاحظ أيضا كما فى المقطع السابق كمية التنسيق بين ما يدخل و ما يخرج عن طريق بوابات و الات نقل متخصصة فالمسألة ليست مجرد كائن قرر أن يقضى حياته فى قلب كائن اخر بل هناك درجة عالية من التكامل و الاندماج بينهما و هذا المستوى من الاندماج لا يمكن تفسيره بالنشوء التعايشى حيث يحتاج الى الاتى:
1-يخسر الكئن الدخيل معظم جيناته لأنها غير موجودة فى العضية
2-يطور جينات جديدة للوظيفة الشاغرة فى الكائن المضيف
3-تطور الخلية المضيفة معظم الجينات التى يحتاجها الكائن الدخيل مرحبة به
4-تتطور اليات للتبادل بين الكائنين
و كمثال بسيط فان الميتوكوندريا تستورد الكثير من احتياجاتها من الخلية المضيفة و لأجل هذا فان البروتينات الموجهة نحو الميتوكوندريا يتم وسمها بتسلسل خاصة يعمل كاشارة توجيه لكى يتم نقل هذه الى الميتوكوندريا تحديدا و منها جهة أخرى تحتوى الميتوكوندريا على بوابات متخصصة فى غشاءها الخارجى translocase of the outer membrane TOM و كذلك فى غشاءها الداخلى translocase of the inner membrane TIM للتعرف على هذه البروتينات و تمريرها الى الداخل فى ظل حماية من بروتينات أخرى متخصصة تسمى الشابيرونات تمنع الشحنة من بدء التطوى و اتخاذ الشكل ثلاثى الأبعاد أثناء النقل قبل الدخول الى الميتوكوندريا حتى لا يعيقها ذلك عن المرور من البوابات. أما ان كان البروتين لن يدخل الى قلب الميتوكوندريا بل سيعمل فى منطقة الغشاء فهذا أيضا له اشارة خاصة يتعرف عليها مركب بروتينى ثالث يدعى OXA ليضعه فى مكانه فى الغشاء الداخلى و مركب SAM ليضع البروتينات فى الغشاء الخارجى و طبعا هناك بروتينات signal peptidase التى تزيل الوسم لاحقا عن البروتين بعد أن يصل الى وجهته حتى لا يتداخل مع عمله. كل هذه الأجزاء من اشارات و بوابات يجب أن تنشأ هكذا فجأة حتى يكون لعملية الابتلاع المزعومة أى فائدة تكيفية و الا فان القائم بها لن ينتشر فى الجماعة الحية
The origin of the machinery for protein import is more complicated and is subject to much debate...Most of the transferred genes continue to support mitochondrial functions, having somehow acquired the targeting sequences that allow their protein products to be recognized by TOM and TIM and imported into the organelle.
Franklin Harold, In Search of Cell History: The Evolution of Life’s Building Blocks (Chicago: University of Chicago Press, 2014), 137-138
أضف الى كل ذلك أن كود تشفير الميتوكوندريا يختلف عن البكتيريا [مما ينفى كون أصلها نشوء تعايشى] و عن الخلية حقيقية النواة [مما ينفى كونها نشوء تطورى ذاتى]
Darnel, James, Harvey Lodish and David Baltimore. 1986. Molecular Cell Biology. New York: Freeman
الطريف أنه فى وسط كل هذا لم تحل النظرية المشكلة بل أخذتها خطوة الى الخلف و شوشت عليها باغراقها فى فيضان من التفاصيل المفترضة فالمشكلة الأصلية هى أن حقيقيات النواة تمتلك الية معقدة لانتاج الطاقة اسمها ميتوكندريا لا سلف لها فبدائيات النوى لا تملكها و لا توجد أى أشكال وسيطة أو مراحل تطورية اذن من أين أتت بها؟
تقول النظرية باختصار أتت بها من خلية أخرى دخيلة invading cell حقا انه تفسير رائع و منطقى لكن من أين أتت بها الخلية الدخيلة ابتداءا و من أين امتلكت هذا النظام الوظيفى الصناعى المعقد؟؟؟ ان جوهر فكرة النشوء التعايشى هو انضمام زوجا من الخلايا أو الأنظمة المنفصلة [أى أنها تفسر وجود الشئ بأنه موجود!!!] دعونا نلقى نظرة مثلا على المحرك البروتينى المعقد المغروس فى جدران الميتكوندريا ATP Synthase و الذى يعمل كروبوت الى على خط انتاج يقوم بمعالجة و تركيب أجزاء الجزيئات لانتاج منتج نهائى

و لنتجاهل قليلا العقبات التى تواجه الفرضية و نتظاهر أننا نصدق أنه جاءها من الخارج...من أين جاء هذا التعقيد الوظيفى الصناعى؟ بالتأكيد ليس من التراص العشوائى للجزيئات.
ان بدائيات النوى لا تحتوى على عضيات مثل الميتوكوندريا لذلك فهى تقوم بعملية معقدة لتصنيع ال ATP عالى الطاقة فى الغشاء عن طريق هيكل فريد مصمم لهذا الغرض لا يوجد أى مماثل له فى حقيقيات النوى و لا فى الميتوكوندريا ذاتها مما ينفى أن هذه الخلايا هى أصل الميتوكوندريا و هذا واحد فقط من العديد من الفروق بين الميتوكوندريا و البكتيريا السلف المزعومة بل ان حتى بعض حقيقيات النوى الخالية من الميتوكوندريا و التى افترض التطوريون أنها سلف لحقيقيات النوى الحالية ذات العضيات أو نسل لحقيقيات النوى العتيقة قبل أن تكتسب العضيات اتضح احتواء جينومها على جينات ميتكوندريا مما ينفى كونها سلفا أو امتدادا لخلايا ما قبل العضية
فى النباتات تقوم البلاستيدات الخضراء chloroplasts بانتاج جزئ الطاقة ATP باستخدام الكلوروفيا [اليخضور] بينما تستخدم بكتيريا التمثيل الضوئى [السلف التعايشى المفترض للبلاستيدات] نظاما اخرا تماما غير البلاستيدات و بالتالى مرة أخرى توجد فجوة تطورية عملاقة بلا أشكال انتقالية تدحض النشوء التعايشى
فى النهاية بقى أن نقول أن عملية تصنيع ال ATP من المواد الغذائية و التعامل معه عملية حيوية كيميائية غاية فى التعقيد سواءا فى حقيقيات النواة أو بدائيات النواة بشكل لا ينتج الا عن تصميم و علم و قصد و غاية
تطور العضيات:
اذا تركنا النشوء التعايشى قليلا و عدنا الى التطور الكلاسيكى فان تطبيقه على العضيات مستحيل ليس فقط لغياب الأشكال الانتقالية أو لاختلاف الاليات المستخدمة بين بدائيات النوى و عضيات حقيقيات النوى أو لاستحالة الاندماج بهذا الشكل و لكن أيضا بسبب مفهوم التعقيد غير القابل للاختزال irreducible complexity الذى ناقشناه فى فصلين سابقين. العضيات هياكل معقدة يتألف كل واحد منها بدوره من الاف الأجزاء المعقدة بدورها فمثلا لا تتجول البروتينات بمفردها فى الخلية بعد تصنيعها بل تستخدم اليات نقل معقدة منها نظام البوابات الذى يحتاج الى انشاء البوابة و انشاء الية داخل غشاء الخلية للتحكم فيها و فتحها و انشاء جهاز استشعار كيميائى يرصد اقتراب البروتين المطلوب من البوابة لينشط الية الفتح و كل جزء من هذه الأجزاء بدوره مركب من أجزاء معقدة يجب وجودها كلها معا بشكل متزامن و قابليتها للاندماج و العمل معا و أمثلة مثل هذه الاليات كثيرة جدا
Robert H. Singer, "RNA zipcodes for cytoplasmic addresses,” Current Biology 3 (1993): 719—721. doi:10.1016/0960-9822(93)90079-4. PMID:15335871.
Donald M. Engelman, "Membranes are more mosaic than fluid,” Nature 438 (2005): 578—580. doi:10.1038/nature04394. PMID:16319876.
Jonathan Wells, "Membrane patterns carry ontogenetic information that is specified independendy of DNA,” Bio-Complexity 2 (2014): 1-28. doi:10.5048/BIO-C.2014.2.
أصل النواة:
يفترض التطوريون أيضا أن نواة الخلية ذاتها جاءت عن طريق النشوء التعايشى [يلا هى جت على دى] حيث أن المشكلة الرئيسية فى نشأة حقيقيات النوى هى النواة ذاتها قبل العضيات فيفترض الدراونة أن جرثومة عتيقة قد اندمجت مع بكتيريا و أصبحت نواتها [ما هى زاطت بقى] المشكلة أن هذا يتطلب امتلاك الخلية المضيفة تركيبا معقدا يسمح لها بالابتلاع phagocytosis و ألا تمتلك جدار خلوى بينما مواصفات بدائيات النوى عكس ذلك تماما فهى بلا هيكل خلوى معقد و تمتلك جدار خلوى لذا فهى غير قادرة على اتمام العملية
Hartman, Hayman and Alexei Fedorov. 2002. "The Origin of the Eukaryotic Cell: A Genomic Investigation". PNAS. 99(3):1420
اضافة الى أن التعقيد التنظيمى لحقيقيات النوى أكبر بكثيرمن بدائياتها مما يصعب جدا تصور نشأة النوى من بدائيات النوى
Hickman, Cleve, Larry Roberts and Alan Larson. 1997. The Biology of animals WCB-McGraw Hill. p.39
بل لقد ذهب بعض الدارسين الى كون الخلية الأولى كانت هى حقيقية النواة ثم خسرت النواة بعد ذلك و هذا هو عكس نظرية النشوء التعايشى و قد كان أحد دوافع وضع هذا النموذج هو أن مزاعم الاتساق الجينى للأشجار التطورية لها معارضات عديدة اذ أن التشابهات التى تستدل بها دائما نظرية التطور - بغض النظر عن أنها ليست دليلا أصلا- تنتج أشجار و علاقات تطورية مختلفة و متضاربة و ليست متسقة فبعض الجينات تبدو قريبة الى نوع و البعض الاخر الى نوع اخر و هكذا
Patrick Forterre and Herve Philippe, “The Last Universal Common Ancestor (LUCA), Simple or Complex?” Biological Bulletin 196 (1999): 373—377.
Patrick Forterre and Herve Philippe, “Where Is the Root of the Universal Tree of Life,” BioEssays 21 (1999): 871—879.
و لعل من الأمثلة المضحكة على هذا أن رسم شجرة تطور الحيتان-التى يفترض كونها حاليا واحدة من أقوى الأمثلة التطورية- عن طريق جينات الميتوكوندريا قد تضارب مع رسمها عن طريق السمات التشريحية
morphologists have classified the cetaceans and mesonychids together as a sister group to the Artiodactyla... mitochondria, which placed cetaceans in the middle of the Artiodactyla.
Trisha Gura "Bones, molecules…or both?" Nature volume 406, pages230–233 (2000)
فبدلا من أن تؤكد لنا الميتوكوندريا السلف المشترك طعنت فيه و ليست شجرة الحيتان فقط بل ان محاولة رسم شجرة تطور للمايتوكوندريا بشكل عام أدى الى نتائج تتعارض مع الفرضية الأساسية لشجرة الحياة

اليوم العالمى للمكرونة الاسباجيتى - أقصد شجرة تطور المايتوكوندريا
Vicky Merhej and Didier Raoult "Rhizome of life, catastrophes, sequence exchanges, gene creations, and giant viruses: how microbial genomics challenges Darwin" REVIEW article Front. Cell. Infect. Microbiol., 28 August 2012 Sec. Molecular Bacterial Pathogenesis
لا يسعك و أنت تشاهد هذا العبث الا أن تتذكر رتشارد دوكنز و هو يؤكد للناس أن أحد أقوى و أهم أدلة التطور هو امكان تنظيم الجينات و الصفات التشريحية على هيئة أشجار متسقة - مجموعات ضمن مجموعات nested hierarchies فى شكل هرمى أو شجرى متفرع و هو توصيف لا علاقة له بكومة النباتات المتسلقة أعلاه. الميتوكوندريا التى من المفترض أنها من أدلة السلف المشترك بلا سلف مشترك!!! هل أخذت الخلية البدائية تتبضع بين الأنواع المختلفة لتأخذ بضع جينات من نوع و بضع من نوع اخر و هى تبتلع شتى انواع الخلايا حولها ثم يقوم هؤلاء بارسال جيناتهم الى نواتها بكل أريحية لتنتقى و تختار ثم تتقاذف الخلايا جينات الميتوكوندريا فيما بينها حتى تصنع لنا نموذج كهذا؟ بعض التطوريين يقول هذا و يزعم أن السلف المشترك كان عبارة عن مجموعة خلايا بجينات مختلفة تتبادلها مع بعضها البعض و هذا طبعا ينقل المشكلة فقط الى مكان اخر و هو نشأة تلك الخلايا ذات الجينات المتباينة. هل حدث انتقال أفقى للجينات بكثافة شديدة عبر تاريخ تطور الكائنات و أخذت الكائنات تتبادل الجينات عن طريق عمليات عدوى فيروسية أو بكتيرية؟ بعضهم أيضا يقول هذا فى محاولة لتبرير تضارب البيانات. المشكلة هنا أن كل هذه سيناريوهات اضافية لتبرير لماذا لا يعمل دليل التطور و الحقيقة الوحيدة المثبتة هى أن الدليل لا يعمل.
مفتش الشرطة: لقد عثرنا على المجرم و لدينا الدليل الدامغ - بصماته تملأ المكان
القاضى: يا سيادة المفتش - أين البصمات التى تتحدث عنها؟ انها غير موجودة
مفتش الشرطة: لقد أزالها يا سيدى لذك هى غير موجودة. ألم أقل لك انه مجرم عتيد الاجرام
القاضى: !!!!!!!!!
ان التشابه بمنتهى البساطة تشابه وظيفى بدون الحاجة الى اسباجيتى تبادل الجينات الغير واقعى هذا و الذى يهدف فقط الى الحفاظ على النظرية. و أخيرا و ليس اخرا ففى مراجعة للنظريات المختلفة لتفسير نشاة الميتوكوندريا و العضيات طرح الباحثون مجموعة من الأسئلة التى يجب على النظرية التفسيرية اجابتها و خلصوا الى أنه برغم مئة عام من البحث فان كل النظريات المطروحة لم تتمكن من استيفاء اجابات كل الأسئلة و أننا بحاجة الى نظريات جديدة (لاحظ أنهم بالعقلية التطورية لذا أحيانا يقبلون فرضيات غير مثبتة كاجابات و يعتمدون التشابه كدليل على السلف المشترك متجاهلين أن الكثير من التشابهات بين الميتوكوندريا و أسلافها المرشحين سببه ببساطة الوظيفة و محددات العمل و ضوابط البيئة)
!) unique, singular origin of eukaryotes and mitochondria;
2) lack of intermediate, transitional forms;
3) chimaeric nature of eukaryotes, especially membranes;
4) lack of membrane bioenergetics in the host;
5) lack of photosynthesis in symbiont;
6) origin and present phylogenetic distribution of MROs.
7) the original metabolism of host;
8) the original metabolism of symbiont;
9) the initial ecological relationship of the partners that specified the initial conditions and restrictions of the merger, and what stabilized this relationship;
10) the early selective advantage of the partnership;
11) the mechanism of inclusion;
12) the mechanism of vertical transmission of the proto-endosymbionts.
Istvan Zachar and Eors Szathmary, “Breath-Giving Cooperation: Critical Review of Origin of Mitochondria Hypotheses,” Biology Direct 12 (August 14, 2017): 19
كل هذا يتم تجاهله و يتم الترويج لفرضية النشوء التعايشى على أنها حقيقة مثبتة لمجرد أنها الأشهر
الميتوكوندريا و التصميم:
احدى الدراسات التى أرادت أن تقيم أهمية الميتوكوندريا للحياة وجدت أن الخلايا لديها مقياس مهم جدا اسمه: الطاقة لكل جين Available Energy per Gene (AEG) و الخلايا حقيقية النوى و هى تلك الخلايا ذات درجة التعقيد المطلوبة لبناء المخلوقات المتقدمة متعددة الخلايا لديها تحتاج الى الاف الأضعاف من هذا المقياس مقارنة بالخلايا بدائية النوى كالبكتيريا. هنا يأتى دور الميتوكوندريا. يؤكد لنا التطورى أن الميتوكوندريا بهذا الشكل و بهذه الخصائص لمجرد الصدفة الجميلة التى جعلت خلية تبتلع خلية ثم جعلت الفريسة تفقد جيناتها. على الجانب الاخر تؤكد لنا البيانات (بدون حشر اراء و تفسيرات التطوريين الشخصية) أن الميتوكوندريا بهذا الشكل لتحقق وظيفة مهمة جدا ما كان لها أن تتحقق الا بهذا الشكل لأن اختزال الجينوم و ابقاء فقط ما هو مطلوب لتوليد الطاقة (بناء سلسلة نقل الالكترونات ETC و ضروريات الغشاء الحامل لها) يؤدى الى تخفيض الطاقة المطلوبة بشكل كبير لهذه العضيات مقارنة بخلية كاملة و بالتالى زيادة كفاءتها و معدلات انتاجها. و فى المقابل و بعكس ما يقوله بعض التطوريين و يستدلون به على أنها مجرد فريسة طال عليها الأمد فان الميتوكوندريا ليست فى طور خسارة جيناتها بالكامل بل ما تحتويه من جينات ضرورى و باقى (أرجو هنا أن تلاحظ المنطق الأعرج! كان التطوريون يقولون هى فى مرحلة خسران الجينوم و بعد ملايين السنين ستصبح بلا جينوم و لو كانت مصممة لنقل المصمم جينومها بالكامل الى قلب النواة لتكون عمليات التصنيع و الترجمة مركزية فهذا أفضل و أوفر ثم لما اكتشفوا مزايا للابقاء على بعض أجزاء الجينوم فيها لتسهيل و تحسين عمليات النقل و الانتاج و تسريع الاستجابة للمتغيرات التى قد تتطلب تعديل سريع فى معدل انتاج بعض المكونات الحيوية بعكس ما كانوا يقولون نسبوها للانتخاب الطبيعى كما فى المصدرين الثانى و الثالث أدناه.)
Nick Lane, “Bioenergetic Constraints on the Evolution of Complex Life,” Cold Spring Harbor Perspectives in Biology 6, no. 5 (May 2014): a015982
Iain G. Johnston and Ben P. Williams, “Evolutionary Inference across Eukaryotes Identifies Specific Pressures Favoring Mitochondrial Gene Retention,” Cell Systems 2 (February 2016): 101–11,
Patrik Björkholm et al., “Mitochondrial Genomes Are Retained by Selective Constraints on Protein Targeting,” Proceedings of the National Academy of Sciences, USA (June 2015), doi:10.1073/pnas.1421372112.
وجدت الدراسة الأولى أنه بدون هذه العضيات المتخصصة ذات الجينوم المصغر فان كلفة انتاج الطاقة على أى خلية بكتيرية تخضع لزيادة التعقيد ستكون عالية جدا لدرجة أنها ستبتلع انتاج الطاقة الاضافى بل ربما ما هو أكثر. للتبسيط تخيل مثللا محطة توليد طاقة سنضع فيها ماكينات اضافية تزيد من انتاجها بنسبة 625%. المشكلة أن الماكينات الاضافية لكى تعمل تحتاج الى استهلاك طاقة أكبر مما تستهلكه المحطة الان بأكثر من 625% لدرجة أن اجمالى الطاقة سينقص بنسبة 25% بسبب شراهة استهلاك الماكينات الجديدة. واضح جدا أن توليد الطاقة بهذه الطريقة كارثة و سيؤدى الى الفشل. فقط تصميم صغير لماكينات مصغرة تم حذف الكثير من محتوياتها الغير ضرورية لتوليد الطاقة و لكنها كانت ضرورية لوظائف أخرى فى المحطة هى التى ستقوم بالعمل. الان لدينا ماكينات مصغرة للطاقة تستهلك أقل لأنها لا تفعل شيئا سوى توليد الطاقة الى جانب الماكينات الأصلية التى كانت تقوم بأدوار هامة اضافية غير توليد الطاقة لحفظ المحطة ككل. المحطة القديمة قبل التحديث هى الخلية بدائية النوى و الجديدة بعد التحديث هى حقيقية النوى و الالات الجديدة المبتكرة التى تم حذف وظائفها الغير ضرورية هى الميتوكندريا.
طبعا صاحب الورقة أعلاه تطورى و يحاول حشر تفسيره الصدفوى للأمر و كأن هذه الصدفة و ما تبعها من صدف أدى الى تطور باقى عضيات الخلية هى التفسير المنطقى و لكن كما نرى هناك تفسير أفضل بكثير من وجهة نظر التصميم. يخبرنا التطوريون أن القول بالتصميم غير علمى و يتاقض نصل أوكام و مبدأ الاقتصاد فى الفرضيات parsimony و لكن ما يفعلوه هو ازاحة فرضية التصميم برغم دعم البيانات لها ثم وضع عشرات الفرضيات بدلا منها متمثلة فى صدف متتالية لابتلاع خلية أخرى ثم فقد الفريسة للجينوم أو انتقاله للخلية العائلة و تطويرها لاليات تضخ بها بطاريات الطاقة الاضافية ATP لعائلها ثم خضوعها لعملية تعديل فى الشفرة الجينية (شفرة الميتوكوندريا فيها اختلافات عن شفرة الخلية) و خضوعها أيضا لعمليات تبادل جينية مع كائنات أخرى للتغطية على السلف المشترك! بل ان التطورى و هو يحدثك عن الاقتصاد فى الفرضيات لن يذكر لك مثلا أن بعض فرضياته تتطلب دخول البكتيريا الى الخلية ثم هروبها منها بعد فترة حتى ينشأ منها خطان تطوريان منفصلان أو دخولها مع بكتيريا أخرى تدعمها و تساعدها على اجتياز الدفاعات و المناعة...ما شاء الله على الاقتصاد فى الفرضيات و المعقولية الرهيبة بدلا من خرافات التصميم التى يسمونها سحرا
Living cyanobacteria neither possess the genetic toolkit to evade host defenses, nor do they encode effector proteins to interact with the host cellular machinery. So, how did the unprotected photosynthetic cell survive the early phases of the endosymbiosis?...chlamydial bacterium entered the host cell together with a cyanobacterium. This allowed the cyanobacterium to escape host defenses and establish a tripartite symbiosis through the help of chlamydial-encoded effector proteins and transporters. However, the details of this complex process remain incompletely understood.
Ball, S. G., Bhattacharya, D. & Weber, A. P. M. Pathogen to powerhouse. Science 351, 659–660 (2016).
We note, however, that the endosymbiont did not have to be an obligate intracellular bacterium at the time of the initial endosymbiosis event. As a result, it could have escaped the host later on and given rise to obligate intracellular Rickettsiales lineages as we see today
Wang, Z. & Wu, M. An integrated phylogenomic approach toward pinpointing the origin of mitochondria. Sci. Rep. 5, 7949 (2015).
An independent-endosymbioses scenario is more probable than the shared-endosymbiosis scenario: the latter requires the unlikely event of a hypothetical endosymbiotic ancestor of Rickettsiales and mitochondria escaping the host cell before engaging a new endosymbiosis that gives rise to the Rickettsiales
Joran Martijn et al., “Deep Mitochondrial Origin Outside the Sampled Alphaproteobacteria” Nature 557 (May 3, 2018): 101–5
هل أدركت ما الذى يحاولون فعله؟ عندما يقوم العلماء بزراعة عضو جديد فى الجسد فأحيانا يضطرون الى حقنه بمضادات لكبت الجهاز المناعى حتى لا يهاجم العضو الجديد. هؤاء حرفيا يحاولون نسبة نفس العملية لعمليات عشوائية طبيعانية فقط لتجنب القول بالتصميم. ناهيك طبعا عن عمليات الابتلاع المتعددة المطلوبة لانتاج مختلف العضيات فى قلب الخلية غير الميتوكوندريا و ما يصاحبها من تطور اليات التكافل بين البالع و المبلوع كبروتينات OXA/SAM/TIM/TOM المذكورة أعلاه و ما يصاحبها من اشارات بأخطاء فى نسخ احمض النووى...منتهى الاقتصاد فى الفرضيات و التفسيرات العقلانية. و هذه الدراسات بالمناسبة لها دلالة أخرى هامة غير كونها اشارة الى وجود تصميم. ان الكثير من فرضيات النشوء التعايشى تحتاج الى أن تطور البكتيريا تعقيدا يمكنها من الوصول الى مرحلة ابتلاع خلية أخرى و هو ما يسمى أحينانا بمرحلة انتقالية نحو حقيقيات النوى أو حقيقيات نوى بدائية و لكن ان كان تطوير التعقيد يتطلب الميتوكوندريا لتوليد طاقة داعمة لتعقيد و الميتوكوندريا تتطلب التعقيد حتى تتمكن الخلية من بلع زميلاتها فاننا الان نعانى من حالة من حالات البيضة و الدجاجة. الميتوكوندريا تحتاج تعقيد و التعقيد يحتاج ميتوكوندريا.
الخلاصة:
يقول البيولوجي (ألبرت دي روز)، والذي لا يتبع الخلقوية ولا التصميم:
"حينما نحاول أن نقحم سيناريو ميكانيكي بناء على نظرية التعايش الجواني endosymbiotic theory فسريعا ما نواجه المشاكل. الطفرات الجينية التي ستسمح للبكتيريا بالازدهار في السيتوبلازم (العصارة الخلوية) لن تكون عملية للبقاء على المدى الطويل. الخلايا اللاهوائية عادة لا تنجو في بيئة تحتوي على الأكسجين، بينما الـمُعايش الـجَوّاني endosymbiont سيحتاج إلى الأكسجين لكي يعكس أفضلية في الكفاءة. كلا الكائنين في البداية سيتنافسان على مصادر الطاقة؛ حيث أن البكتيريا مستهلكة لأدينوسين ثلاثي الفوسفات ATP وليست مصدرة له. الانتقال المكثف للجينات الذي تطلبه النظرية سيعيث فسادا في جينوم معقد، ذلك لأن الإدخال المتكرر لقطع عشوائية من دنا الميتوكوندريا mtDNA سيعطل الوظائف الموجودة. علاوة على ذلك، انتقال الجينات هي عملية متعددة الخطوات، حيث يجب أن تنتقل الجينات فيها إلى النواة، وأن يتم التحايل على الاختلافات الجينية للميتوكوندريا، وأن تتم عملية التعبير الجيني بشكل صحيح، وأن تعاد أيضا إلى الميتوكوندريا لكي تكون وظيفية.اختصارا؛ السيناريوهات الآلية لنظرية التعايش الجواني تتضمن أشكالا وسيطة غير وظيفية أو ضارة تماما للكائن".
-نقلا عن مركز براهين
خلص التطوريون الى أن "الميتوكوندريا كانت حادثا محظوظا [و ما أكثرها فى التطور] أولا اكتسبت خلية سلف قديمة القدرة على ابتلاع و هضم جزيئات معقدة [و احنا قاعدين] ثم بدأت تفترس أقرانها من المايكروبات [و احنا قاعدين] ثم فى مرحلة ما توقفت عن هضم فرائسها بالكامل [و احنا قاعدين] ثم تناولت وجبة أقامت بداخلها [و احنا قاعدين] ثم تطورت هذه الوجبة السعيدة لتصبح الميتوكوندريا [فاحنا اتخضينا بقى!!!] " و طبعا تكرر هذا السيناريو عدة مرات لتفسير عضيات أخرى. و تتبقى أمام هذه القصة اللطيفةعقبة أخيرة: دراسة جينات حقيقيات النوى تشير الى أنها احتوت على الميتوكوندريا من قبل أن تكتسب القدرة على ابتلاع الخلايا الأخرى.
ان التشابه بين السيانوبكتيريا و الكلوروبلاست ببساطة لأن كلاهما يقوم بوظيفة التمثيل الضوئى و نفس الأمر بالنسبة للميتوكوندريا و البكنيريا...التشابه وظيفى و ليس لأن كائنا ابتلع اخر و دمجه فى نفسه تماما كما أن التشابه بين معالج البيانات processor فى جهاز كمبيوتر و اخر تشابه وظيفى و ليس لأن أحدهما ابتلع الاخر. أضف الى ذلك أن العلاقات التكافلية شائعة بين الكائنات الحية فهناك بكتيريا تعيش فى أمعاء الانسان و أخرى تتبادل المنافع مع جذور النباتات و فطريات تعيش فى مستعمرات النمل بل و أسماك تنظف أسنان أسماك أخرى و طيور تنظف أسنان التماسيح. فى كل حالة من هذه و غيرها الكثير هناك تبادل منفعة بين الكائنات و أحيانا حياة مشتركة كحالة البكتيريا و الانسان و لكن هذا ليس معناه أننا ان تركناهم فسيندمجون فى كائن واحد لنجد بعد مليون سنة انسانا له سوط بكتيرى!!
و أخيرا وجب التنبيه على نقطة فى منهى الأهمية...ان المحور الرئيسى فى نظرية لين مارجوليس هو الحصول على جينومات موجودة بالفعل...فالخلية المعقدة حصلت على جينومات من خلايا أبسط و الكائنات المعقدة حصلت على جينوامت من كائنات أخرى عن طريق الانتقال الأفقى للجينات الا أن هذه النظرية تعتمد على وجود كل المكونات ابتداءا ثم مزجها و تركيبها بطرق مختلفة لانتاج الكائنات الحية و لكنها لا تفصل نشأة هذه المكونات نفسها الا بالحشو الكلامى المعتاد عن الطفرات و الانتخاب أى أننا حتى اذا افترضنا صحة هذه النظرية تنزلا فاننا لم نفسر شيئا فنصبح كمن يقول الكتب موجودة لأن الجمل التى صنعت منها الكتب تم تركيبها معا لصناعة هذه الكتب!!!

منقول من ملخص كتاب "تفنيد أركان الداروينية" بقلم جيري بيرجمان




تعليق